RENR 690 Jacob Beauregard
Results
and
Discussion

Figure 19. Sugar maple (Acer saccharum) canopy at Mont-Mégantic national park, Quebec (source: Jacob Beauregard)
01
Dominant forest mycorrhizal type is an important driver of fungal community composition
Key Research Findings
02
Effects of dominant forest mycorrhizal type are associated with fungal community structure across host species and compartments (root, soil)
Dominant mycorrhizal type does not have a significant association with pathogen relevant abundances, nor does it associate with pathogen abundances to reduce maple growth
03
Forest dominant mycorrhizal type and fungal community composition
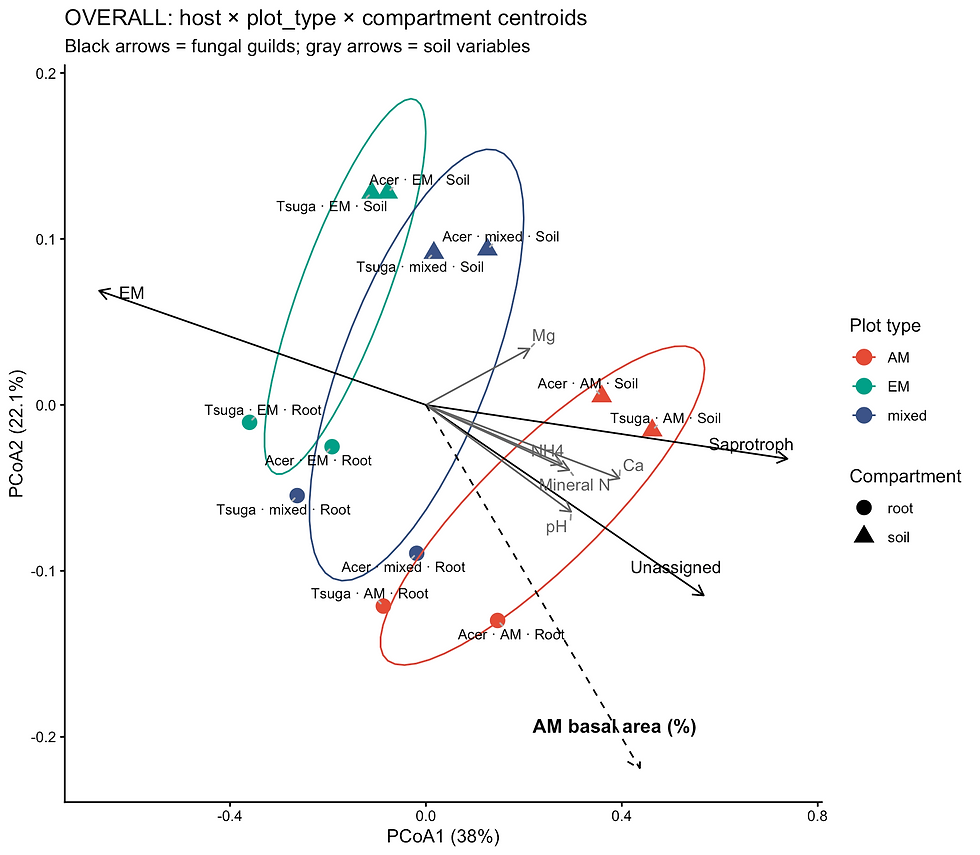
Figure 20 Genus-level PCoA (Bray-Curtis) for fungal communities of different plot types (red = AM; green = EM; blue = mixed) when considering host (Acer saccharum, Tsuga canadensis) and compartment (root, soil) with envfit vectors for guilds and soil chemistry where significant (p<0.05)
Bray–Curtis PERMANOVA revealed significant differences in fungal community composition among forest plot types in both roots (R² = 0.31, p = 0.016) and soils (R² = 0.50, p = 0.031), after accounting for site effects. Root and soil communities also differed significantly within plots (R² = 0.17, p = 0.001). Tests of multivariate dispersion were not significant, indicating that these differences reflect shifts in community composition rather than differences in dispersion.
Principal coordinate analysis (Figure 20) showed clear separation of AM-, EM-, and mixed-dominated plots along the primary axis, reflecting a gradient from AM- to EM-dominated stands, with mixed plots occupying intermediate positions. The first two axes explained 60.1% of the variation (PCoA1 = 38.0%, PCoA2 = 22.1%). This pattern was consistent across host species and compartments, indicating a strong and pervasive effect of forest mycorrhizal type on fungal community structure.
Environmental fitting identified mineral nitrogen (NO₃⁻, NH₄⁺), calcium, and total mineral nitrogen as significant correlates of community structure, with NO₃⁻ showing the strongest association (r² = 0.60, p = 0.001). These variables aligned with the primary gradient observed in the ordination, suggesting that shifts in soil nutrient availability contribute to differences among plot types.
Functional guild composition was also strongly associated with this gradient. Ectomycorrhizal fungi were aligned with EM-dominated plots (r² = 0.72, p = 0.001), whereas saprotrophic, pathogenic, and unassigned fungi were associated primarily with AM and mixed plots. Together, these results indicate that forest mycorrhizal type structures fungal communities through both abiotic (soil chemistry) and biotic (guild composition) pathways.
When host identity (Acer and Tsuga) was incorporated into the analysis, species-specific shifts were observed within plot-type groupings; however, host centroids remained largely within plot-type clusters. This suggests that host identity modifies fungal community composition but does not override the dominant influence of forest mycorrhizal type.
Overall, these results demonstrate that forest mycorrhizal type is a primary driver of fungal community composition in both roots and soils, with consistent effects across compartments and strong alignment between compositional, environmental, and functional patterns.
Forest dominant mycorrhizal type does not have a significant association with maple growth outcomes

Annual growth differed modestly among forest types, with higher median growth in EM-dominated stands relative to AM and mixed stands (when considered by % AM tree basal area)(Figure 21). Substantial overlap in growth distributions was observed among plot types.
These results point to the notion that seedling growth is influenced by multiple interacting factors rather than a single dominant driver, and that forest mycorrhizal type does not have a significant association with maple growth (p = 0.266).
Figure 21. Relationship between sugar maple seedling growth and the proportion of AM basal area across plots. Points represent individual seedlings, and the line shows predictions from a linear mixed-effects model with site included as a random effect (shaded area = 95% confidence interval). No significant relationship was observed between AM basal area and seedling growth (p = 0.266, R²ₘ = 0.01), indicating that variation in forest mycorrhizal composition did not strongly predict growth.
Forest dominant mycorrhizal type does not have a significant association with pathogen relative abundances and does not have a negative influence on growth outcomes


Figure 22. Relationship between sugar maple seedling growth and root pathogen abundance across forest mycorrhizal types (EM-, mixed-, and AM-dominated stands). Points represent individual seedlings, and lines show predictions from linear mixed-effects models with age and elevation held constant (shaded areas = 95% confidence intervals).
Figure 23. Mean pathogen relative abundance (z-scored) across forest mycorrhizal types (EM-dominated, mixed, and AM-dominated stands), separated by sample compartment (root vs soil). Points represent estimated marginal means with 95% confidence intervals.
Linear mixed-effects models revealed no significant main effects of plot type, compartment, seedling age, or elevation on mean pathogen relative abundance. A marginal interaction between plot type and compartment was observed (p ≈ 0.07), suggesting context-dependent variation. Estimated marginal means showed weak and variable differences in pathogen abundance among forest types and between roots and soils, with wide confidence intervals (Figure 23).
When relating pathogen abundance to seedling growth (Figure 22), forest mycorrhizal type-specific trends were observed. Growth tended to increase with pathogen abundance in EM-dominated plots and decline in AM-dominated plots, while mixed plots showed weak or inconsistent relationships. However, effect sizes were small and confidence intervals were wide, and none of these relationships were statistically significant.
Overall, pathogen abundance was highly heterogeneous and did not exhibit strong or consistent relationships with seedling growth across forest types
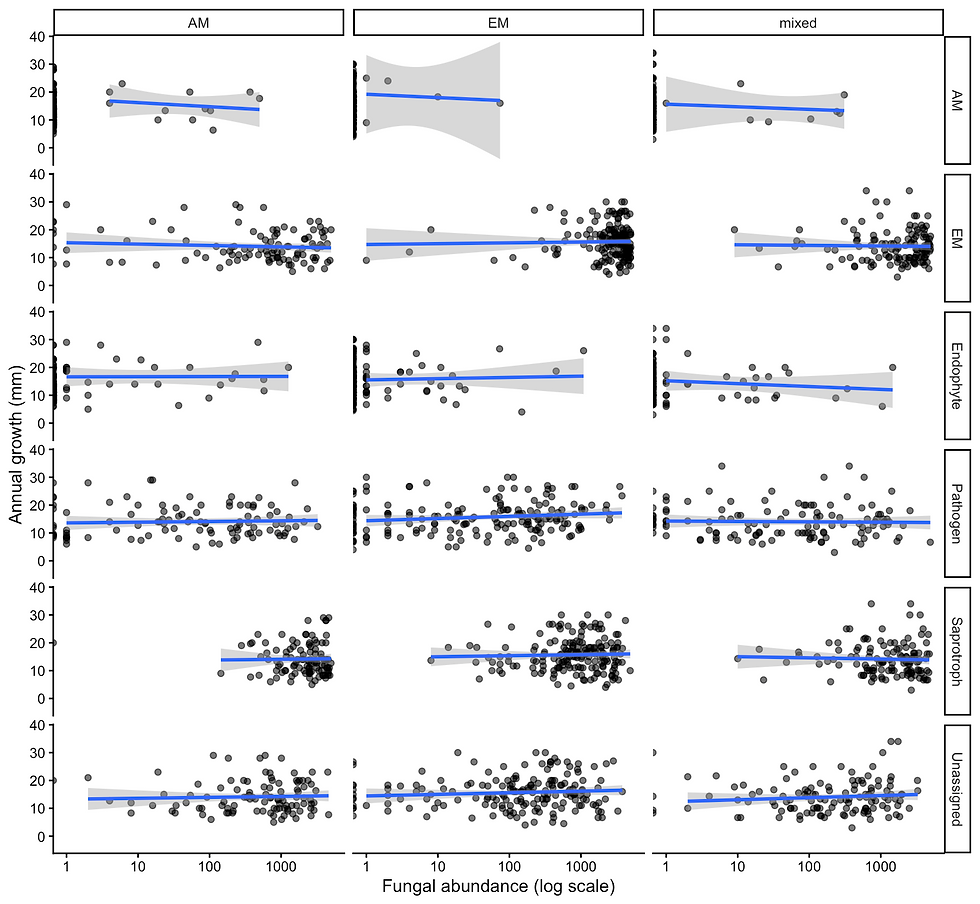
Figure 24. Relationships between fungal functional group abundance and annual seedling growth across forest mycorrhizal types. Points represent individual seedlings, with abundance shown on a log scale. Panels are organized by fungal guild (rows) and plot type (columns: AM-, EM-, and mixed-dominated stands). Lines represent linear model fits with 95% confidence intervals.
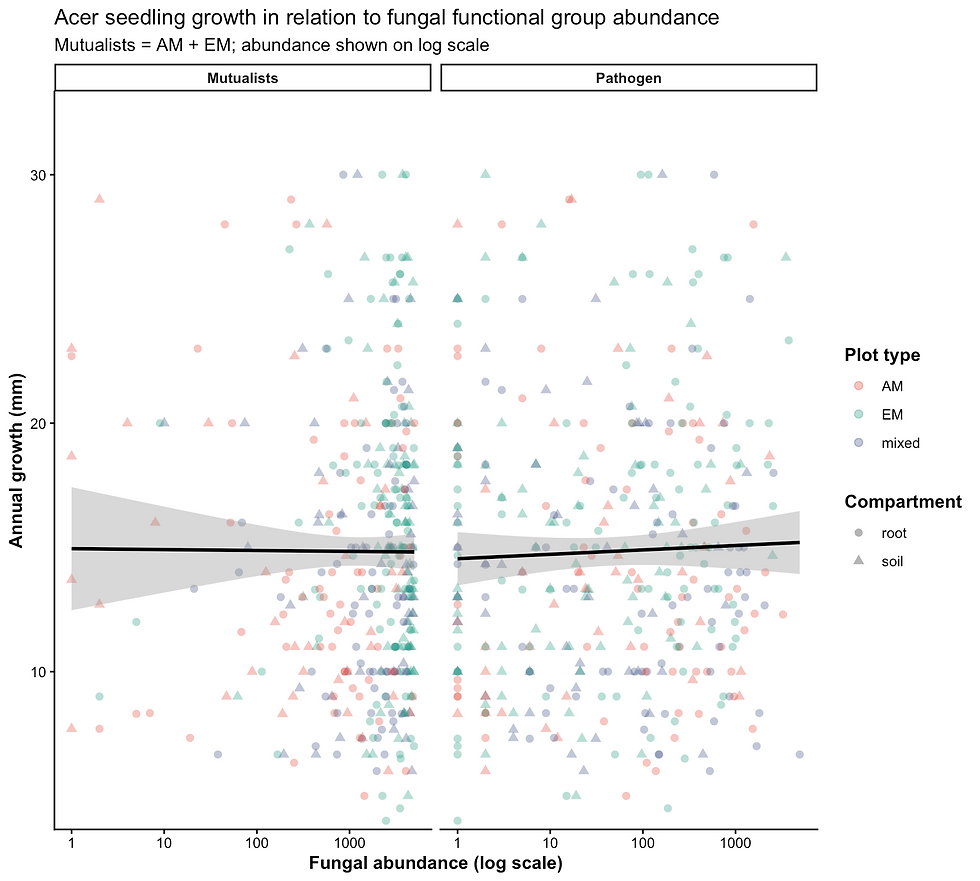
Figure 25. Relationships between seedling growth and combined mutualist (AM + EM) and pathogen abundance. Points represent individual samples, colored by plot type and shaped by compartment (root vs soil), with abundance shown on a log scale. Lines indicate overall linear trends with 95% confidence intervals.
Fungal functional group abundances were not significantly associated with annual seedling growth across any guild examined (all p > 0.3). Similarly, forest plot type did not significantly influence growth within any guild, and no significant interactions between fungal abundance and plot type were detected (all p > 0.1) (Figure 24; Figure 25).
Although visualizations suggested weak variation in abundance–growth relationships among plot types for certain groups (e.g., endophytes), (Figure 24) these patterns were inconsistent and not statistically supported. Across all guilds, relationships between fungal abundance and growth were highly variable, with no clear directional trends. Even when grouping all mutualists together, these relationships remained insignificant (Figure 25)
Together, these results indicate that fungal functional group abundance does not strongly predict sugar maple seedling growth, nor do these relationships differ meaningfully across forest mycorrhizal types.
Discussion
The results of this study revealed that dominant forest mycorrhizal type is a strong driver of fungal community composition in temperate forests, structuring both taxonomic and functional patterns in roots and soils. Fungal communities were organized along a clear gradient from AM- to EM-dominated systems, reflecting coordinated shifts in both soil chemistry and fungal guild composition.
This gradient is consistent with established differences in nutrient cycling between AM and EM forests, where AM systems are typically associated with higher mineral nutrient availability and EM systems with more conservative nutrient economies (Phillips et al. 2013). In our study, this pattern was reflected in strong associations between fungal community structure and mineral nitrogen and calcium availability, suggesting that abiotic conditions and mycorrhizal dominance jointly shape belowground communities.
Despite these strong effects on fungal communities, we found limited evidence that variation in mycorrhizal forest type translated into differences in pathogen abundance or seedling performance. Pathogen abundance was highly variable and not consistently associated with forest type, contradicting the expectation that AM-dominated systems promote pathogen accumulation (Eagar et al. 2022). Although AM stands showed trends toward higher nutrient availability and saprotroph abundance, these conditions did not correspond to increased pathogen prevalence, suggesting that pathogen abundances and interactions are influenced by additional ecological factors beyond the broader forest mycorrhizal context. In addition to pathogen abundance, we assessed relationships between broader fungal functional groups and seedling growth. Across all guilds, these relationships were weak, highly variable, and not statistically significant. This suggests that seedling performance is not strongly determined by any single component of the fungal community, but instead reflects the combined influence of multiple interacting biotic and abiotic factors.
Consistent with these patterns, sugar maple growth did not differ significantly among forest types despite pronounced differences in fungal communities and soil chemistry.
From a functional perspective, these results suggest a divergence between community composition and ecosystem function, where large shifts in microbial communities do not necessarily translate into measurable differences in seedling performance. This has important implications for understanding plant–microbe interactions, as it highlights the potential for functional redundancy, compensatory processes, or emergent properties arising from interactions within fungal communities and between those of other microbes.
In the context of assisted migration, these findings suggest that dominant forest mycorrhizal type alone may not represent a major barrier to sugar maple establishment. Although EM-dominated systems differed substantially in both fungal communities and soil chemistry, these differences did not negatively impact seedling growth. This supports the idea that sugar maple may tolerate a broader range of microbial and abiotic conditions than previously assumed.
However, it is important to acknowledge that our study was conducted within the current range of sugar maple, and EM-dominated stands in this system are not direct analogues of boreal forests. As such, inferences regarding assisted migration remain indirect. Future studies incorporating latitudinal gradients or experimental transplants would be necessary to directly test these hypotheses. While sugar maple maple growth is not negatively impacted by EM stands within the core of its range, this does not speak to their capacity to do the same in boreal forests and across latitudinal gradients which warrant further research.
Together, these findings highlight a decoupling between fungal community structure and seedling performance, emphasizing the need to consider both biotic and abiotic factors when interpreting plant–fungal interactions in natural systems.
Conclusions
This study demonstrates that dominant forest mycorrhizal type is a key determinant of fungal community structure in northern temperate forests, shaping both fungal guild composition and soil chemical conditions across hosts and compartments. Despite these strong compositional differences, we found no evidence that variation in mycorrhizal context translated into differences in pathogen abundance or sugar maple seedling growth. None of the fungal guilds examined were significantly associated with growth, suggesting that mycorrhizal forest type alone is insufficient to predict pathogen pressure or seedling performance.
These findings have important implications for assisted migration. While our study does not directly assess sugar maple establishment in boreal systems, it suggests that environments resembling EM-dominated forests do not inherently constrain maple growth. However, caution is warranted, as boreal ecosystems differ substantially in vegetation structure, climate, and soil properties. Future work incorporating broader geographic gradients and longer-term performance metrics will be necessary to fully evaluate establishment potential.
Overall, our results highlight a disconnect between fungal community structure and seedling performance, emphasizing that large shifts in microbial communities do not necessarily translate into predictable ecological outcomes. This underscores the importance of considering multiple interacting biotic and abiotic drivers when forecasting species responses to climate change and range shifts.