RENR 690 Jacob Beauregard
Dominant Forest Mycorrhizal Types
and Maple Establishment:
A Neglected Biotic Filter?

Background & rationale
Figure 3. Sugar maple (Acer saccharum) canopy at Mont-Mégantic national park, Quebec (source: Jacob Beauregard).


Climate change is reshaping ecosystems, with current trajectories predicting further change into the future (IPCC 2022). Forests are particularly vulnerable to these changes, given the slow regeneration time and limited dispersal capacity of trees (Boisvert-Marsh 2014). With climates projected to change to a much faster degree than trees are capable of adapting, there is concern that many tree species may become extirpated or go locally extinct. In North America, many ecologically and commercially important species are projected to migrate northward to match changing climatic envelopes. With this said, many tree species are migrating at a rate slower than projected and than that of climatic changes (Boisvert-Marsh 2022). As such, there is increasing interest in assisted migration programs, wherein trees are planted in environments that are projected to be analogous to those in which they have evolved and adapted, either inside or beyond their current ranges (Twardek et al. 2023). However, trees do not exist in isolation and rely on a suite of microbial partners that facilitate nutrient exchange, stress tolerance, and pathogen defence (Enea and Beauregard et al. 2025). Thus, important questions remain as to the viability of assisted migration programs when they neglect to consider the resident microbial communities that planted individuals will contend with and how these may impact tree regeneration and long term persistence. When microbial communities are considered for migrating species, it is usually in regard to the facilitative nature of microbes in the establishment of invasive species or how invaders are released from the natural enemies in their native ranges (Heger et al. 2024). As such, critical gaps remain in our understanding of the role of microbial communities in either inhibiting or facilitating migration processes of planted and non-invasive species, particularly for long-lived organisms like trees and in relation to different forest types.

Figure 4. Mature sugar maple (left panel), solitary juvenile maple (top-right panel), sugar maple seedlings (bottom right panel) (source: Jacob Beauregard).
In the temperate forests of eastern North America, few species are as central to this question as the sugar maple (Acer saccharum). The sugar maple (Figure 4) is a keystone forest dominant found across much of northeastern North America (Figure 5) and is of critical importance for both forest ecosystems and industries in the region (Horsley et al. 2002; Collin et al. 2018; Rapp et al. 2019). However, the sugar maple has been experiencing pronounced and prolonged declines across its range, with the exact causes uncertain (Juice et al. 2006; Nolet et al. 2018). Furthermore, the maple is projected to migrate into boreal forests with questions surrounding its capacity to establish and persist in these forests, owing to a suite of soil characteristics thought to be unfavourable to the sugar maple (Collin et al. 2018), including drastically different microbial communities, forest species assemblages, and soil characteristics (Carteron et al. 2020).

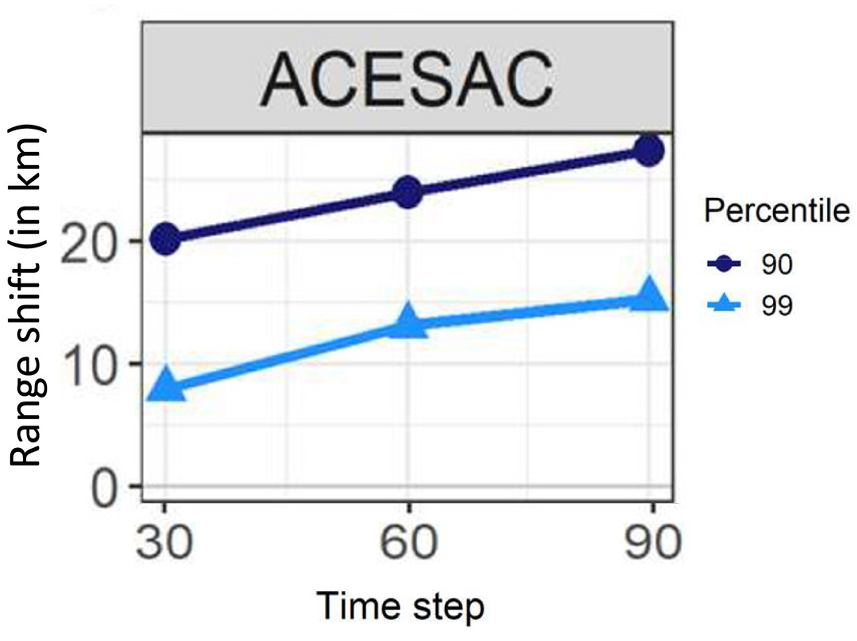
Figure 5. Map of the current distribution for sugar maple (Acer saccharum) in North America (left panel), and future range shifts projections (in km) (right panel). Range shifts are based on the difference between the average latitudinal limit of the simulated distribution as obtained through the simulations in the last year of a given time period (model after 30 years [2011–2040], 60 years [2041–2071], 90 years [2071–2100], respectively) relative to the average latitudinal limit of the initial distribution. At a given percentile (i.e. 90 and 99), positive shifts indicate that the range limit shifted northward between the initial distribution and the time period of interest. (sources: left panel - Enea and Beauregard et al. 2025; right panel - modified from Boisvert-Marsh et al. 2022)
The sugar maple is unique in the high latitude forests of North America in that it is one of the few forest dominants to form obligate associations with arbuscular mycorrhizal (AM) fungi (Cooke et al. 1992). Not only are the majority of temperate tree species ectomycorrhizal (EM), trees in boreal systems are nearly exclusively EM (Collin et al. 2018; Carteron et al. 2022). With this, even if the sugar maple is capable of migrating into boreal systems, questions remain as to its capacity for long-term success owing to a potential lack of critically important symbionts (Chamard et al. 2024).

Figure 6. Graphical depiction of the mycorrhizal associated nutrient economy (MANE) framework, illustrating the soil chemical and biological differences between arbuscular mycorrhizal (AM) and ectomycorrhizal (EM) dominated forests (Phillips et al. 2013).
Furthermore, AM and EM forests have been shown to harbour contrasting soil chemical and nutrient syndromes owing to the characteristics of the dominant trees, the litter they produce, and their fungal partners. Where AM dominated plots are characterized by high levels of available nutrients, EM dominated plots have relatively low levels of available nutrients (Figure 6) with this further contributing to distinct microbial assemblages in the opposing forest types (Phillips et al. 2013). These differing microbial assemblages are proposed to give rise to different plant-soil feedbacks, with feedbacks thought to be more negative in AM-dominated systems, relative to EM systems (Eagar et al. 2025). Specifically, AM forests are thought to harbour relatively higher abundances of pathogenic taxa, owing to their richer nutrient profiles as well as how AM systems and fungi stimulate other microbial taxa. This may contribute to negative frequency-dependence relationships in sugar maple (Bennett et al. 2017; Eagar et al. 2025) and may help to explain the broader regional decline of maple in its core range (Urli et al. 2018; Chamard et al. 2024).


Figure 7. Contrasting resul;tss of seedling transplant experiments, showing survival of planted sugar maple seedlings in the core of their range, at range edge, and beyond at Mont Mégantic, Quebec (source: left panel - Brown and Velland, 2014; right panel - Urli et al. 2018).
While controlled experiments in the region have demonstrated reduced growth of maple due to reduced abundances of root symbionts when grown in soils taken from outside of its core range (Carteron et al. 2020), observational studies and transplant experiments have noted increased growth and survival of maple when grown at its range edge, (Figure 7; left panel) as well as beyond its current range limit (Figure 7; right panel) (Brown and Vellend 2014; Urli et al. 2018; Chamard et al. 2024).
As such, despite the potential for decreased abundances of AM fungi in boreal and EM dominated forest systems, reduced abundances of pathogens and competitive pressure from conspecifics may play a more facilitative than inhibitory role in maple migration. In particular, the ‘enemy release’ hypothesis posits that species may benefit from reduced pressures from species-specific enemies (Urli et al. 2018) and this, coupled with the potentially inhibitory nature of EM dominated stands towards pathogens, may help to explain these varied outcomes in sugar maple growth and establishment through a reduction in pathogen pressure and intraspecific competition. With that, an important gap in our understanding of these varied outcomes in the growth and survival of sugar maple is the potential influence of EM stands on belowground fungal communities and particularly in regard to pathogens. Owing to the overwhelming interest in assisted migration programs for the sugar maple (Zeng et al. 2022), along with its importance to various industries and cultures, it will be important to consider the microbial communities of the forests maple may migrate in to, as these may influence whether maple plantations fail or succeed. To do this, however, it will first be important to assess the performance of sugar maple in EM stands found within its current range. Furthermore, while soil physiochemical characteristics are often considered in this dynamic (Juice et al. 2006), the biotic context is often neglected and dominant mycorrhizal types are largely ignored. Given the potential of dominant forest mycorrhizal types to alter soil chemical regimes and microbial communities, these dyamics should be considered, particularly for species like the sugar maple that will migrate from AM dominated systems to very distinct forest ecosystems which are almost exclusively EM. Of particular interest is whether or not differing dominant mycorrhizal types in the forests of northeastern North America truly lead to differing soil chemical and microbial profiles, and whether this may have meaningful associations with sugar maple regeneration in these forest types.
Here, we aimed to assess the microbial communities of the sugar maple, in both the roots and soils, in forests dominated by differing mycorrhizal types. We also assessed the microbial communities of the eastern hemlock (Tsuga canadensis) as it is a co-occurring tree species with a contrasting mycorrhizal habit (EM), allowing us to be more confident that any potential effects of forest mycorrhizal type were robust to species identity. We hoped to see whether dominant mycorrhizal types were associated with shifts in key fungal guilds, particularly the decomposer (saprotrophic), pathogenic, and mutualistic (AM and EM) fungal guilds as they all play important roles in nutrient cycling and plant health and growth outcomes. Growth outcomes for the sugar maple were also assessed across different forest mycorrhizal types, due to the interest in assisted migration programs for this species and the lack of investigation into growth and fungal communities in EM stands, and the fact that it's age and annual growth can be estimated in the field, unlike the hemlock.
The primary objective of this study was to determine how dominant forest mycorrhizal type structures belowground fungal communities and whether these differences have functional consequences for seedling performance. Specifically, I aimed to (1) characterize variation in fungal community composition across forest types, host species, and compartments (roots vs. soils), (2) assess whether dominant mycorrhizal type is associated with shifts in fungal functional groups, particularly plant pathogens, and (3) evaluate whether variation in fungal community composition and functional groups is linked to sugar maple seedling growth.
A secondary objective was to consider the implications of these patterns for assisted migration under climate change, given that migrating species may encounter novel fungal communities and soil environments.
My hypotheses were that H1. Dominant forest mycorrhizal type structures belowground fungal community composition, with distinct communities associated with AM-, EM-, and mixed-dominated forests, H2. AM-dominated forests will harbor higher relative abundances of plant pathogenic fungi compared to EM-dominated forests, reflecting differences in soil nutrient regimes and microbial interactions and H3. Sugar maple seedling growth will be greater in EM-dominated forests, where reduced pathogen abundance and altered competitive environments may facilitate performance (consistent with the enemy release hypothesis).
I predicted that dominant forest mycorrhizal types would play an important role in structuring belowground fungal communities, in both the soil and roots of both of our species, with AM stands harbouring more distinct communities relative to the EM and mixed. With this, I further predicted that fungal pathogen abundances would be higher in AM dominated systems, relative to EM or mixed systems. Lastly, I predicted that, despite potentially unfavorable soil characteristics, maple growth would be improved in EM dominated forests, potentially owing to reduced pathogen abundances.
In order to meet these objectives, as well as test my hypotheses and related predictions, I worked across the temperate forests of southern Quebec. In total, I collected 435 seedlings across four sites and according to a gradient of dominant forest mycorrhizal types (EM, mixed, AM). I took a variety of growth and environmental metric measurements, performed soil physiochemical analyses, as well as DNA extractions and sequencing for the roots and associated soils of all seedlings collected. By doing this, I hoped to further refine our understanding of the effects of dominant forest mycorrhizal types on fungal communities in northern temperate forests, how these may interact with fungal pathogens, and whether or not these potential differences may be associated with differential seedling growth outcomes. In sum, I hoped to contribute to an improved understanding of what may inhibit or facilitate maple growth in EM systems and to better inform assisted species migration plans, particularly for the iconic sugar maple.